Little
Effect of Training in the Heat on Cycling Performance at Normal Temperature Joanna P
Morrisona, Will G Hopkinsb, Gordon G Sleivertc Sportscience
6, sportsci.org/jour/0201/jpm.htm, 2002 (4278 words)
|
|
Individual
Differences in Acclimation and Performance Training in the heat provides an opportunity to adapt to increased
environmental temperature before performing in a thermally stressful
environment. After heat acclimation
athletes have a lower core temperature at rest (Buono et al., 1998) and during exercise in the heat (Sawka et al., 1983; Armstrong and
Kenney, 1993).
An increase in sweat rate (Nadel et al., 1974), a decrease in sodium content of
sweat (Kirby and Convertino, 1986), a redistribution of sweating (Kirby and Convertino, 1986), and a lowering of the threshold
temperature for initiating the sweating response (Williams et al., 1967) have also been observed after
heat acclimation. There is also an expansion of plasma volume (Senay et al., 1976; Kirby and
Convertino, 1986), which allows for increased blood
flow to the skin to dissipate heat without seriously compromising blood flow
to muscles. Some of these adaptations
probably account for the enhancement of physical performance in the heat that
occurs with heat acclimation (Young et al., 1985; Kirwan et
al., 1987; Armstrong and Maresh, 1991).
Whether these adaptations enhance performance at normal temperature is
unknown. The purpose of this study was
therefore to determine if adaptations gained through heat acclimation enhance
40-km cycle performance in a thermoneutral environment. Ten
highly-trained males volunteered for this study. The subjects were cyclists, triathletes,
and duathletes who had been racing in the top open men’s racing grade for the
previous year. Age, height, and weight
were 25 ± 6 y, 180 ± 5 cm, and 75 ± 7 kg respectively (mean ± SD). Maximum oxygen uptake (VO2max)
was 64.5 ± 3.1 ml·kg-1·min-1,
as determined in an incremental cycling test (150 W initial load, 50 W·min-1
increments) with respiratory equipment and end-point criteria as previously
described (Caird et al., 1999).
The University of Otago Ethics Committee gave ethical approval for
this project, and all subjects provided written informed consent. One subject’s data were discarded due to
non-compliance with the training requirements of the study.
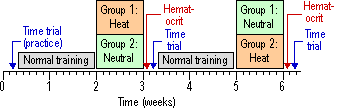
Acclimation Training Acclimation
training was conducted in an environmental chamber set at 37°C and 50% humidity for training in the heat,
or 20°C and 50% humidity for training at
normal temperature. On Day 1 and Day 7
of each acclimation period all athletes cycled on their own bicycles placed
on an air-braked ergometer (Kingcycle Ltd, High Wycombe, Buckinghamshire, UK)
for 90 min at 45% of the subject’s maximum power output attained during the
VO2max test. For the
remaining five days subjects exercised on their own bicycles placed on an
air-braked training device (CS-1000 Cyclosimulator, Cateye, Osaka,
Japan). Each exercise session
consisted of 90 min of self-selected cycle training. The acclimation sessions replaced the
athletes’ training during the seven-day acclimation period. Heart rate was recorded telemetrically
every five minutes and at the end of every work interval (Polar Electro,
Kemplele, Finland). Core temperature
was measured using disposable thermisters (Model 400, Mallinckrodt Medical,
St Louis, MO) inserted 11 cm beyond the anal sphincter. Rectal temperature was sampled every 30 s
on a data logger (1200 Series, Grant Instruments, Cambridge, UK) and recorded
manually every 5 min. Perceived exertion
rated on a 10 point scale (Noble et al., 1983) was recorded every 5 min and at
the end of every work interval. Whole
body sweat rate (ml·h-1) was calculated using measurements of pre-
and post-exercise body mass accurate to 0.01 kg (Wedderburn Scales, Teraoka
Seiko, Tokyo, Japan). Body mass
changes were corrected for fluid consumption, but respiratory water loss and
muscle glycogen loss were assumed to be negligible. Subjects were instructed to dry all sweat
off their body and from their hair before post-exercise mass was
recorded. Work to rest ratios during interval
training were also recorded.
All
subjects completed three 40-km cycling time trials. The first time trial was for
familiarization; the other two were analyzed for the effect of heat
acclimation. Subjects cycled on their
own bikes placed on the Kingcycle ergometer in an environmental chamber. A pre-exercise blood sample, taken from the
subject's left index finger, was collected into heparinised capillary tubes
after the subject had rested quietly on his bicycle for 5 min in his normal
cycling position without cycling. Core
temperature was also recorded at this point via a rectal thermister. The Kingcycle was then calibrated (Palmer et al., 1996) and all participants performed a
10-min self-paced warm-up before starting the time trial. To mimic a competitive event, subjects were
allowed to view a display of speed, elapsed time, and distance covered. Immediately upon finishing the time trial a
post-exercise fingerprick blood sample was obtained from the left index
finger. Three to five capillary tubes
of blood were collected for determination of hematocrit; the mean hematocrit
was used in the comparison of pre- and post-exercise hematocrit to determine
plasma volume shifts during the time trial (Greenleaf et al., 1979).
Subjects were allowed to consume water ad libitum during the time
trial. Whole body sweat rate (ml·h-1)
was calculated from body mass measured before and after the time trial
(accurate to 0.01kg) and corrected for fluid consumption as previously
described. Core temperature, arterial
oxygen saturation (Criticare Pulse Oximeter, Model 504US-504USP, Criticare
Systems, Milwaukee, WI), and heart rate were recorded at 5-km intervals
during the time trial. Mean power
output for the time trial was
obtained from the Kingcycle. Statistical Analyses Changes
in perceived exertion and physiological variables were used as measures of
acclimation. Changes in perceived
exertion and heart rate were calculated by subtracting the mean value over
the last 30 min of exercise during the standard exercise bout on Day 1 from
that on Day 7, then subtracting the resulting neutral acclimation value from
the heat acclimation value. Change in
core temperature was calculated in a similar fashion, but the subject's
resting core temperature (measured immediately before the standard exercise
bout) was first subtracted from the subject's mean value over the last 30 min
of the bout. Change in sweat rate was
calculated by subtracting the sweat rate over the whole exercise bout on Day
1 from that on Day 7, then subtracting the resulting neutral acclimation
value from the heat acclimation value.
Change in hematocrit was the value after heat acclimation minus the
value after neutral acclimation.
Pearson correlation coefficients between measures of acclimation were
used to determine which measures were mutually most consistent. To
determine if heat acclimation changed the physiology of the time trial
performance at room temperature, data from the performance trials were used
to create difference scores for the mean change in core temperature, arterial
oxygen saturation, plasma volume shifts, and heart rate between the heat
condition and the neutral condition.
In all cases data for the neutral condition were subtracted from those
of the heat condition. We
quantified the effect of acclimation on time-trial performance with a
repeated-measures analysis using Proc Mixed in the Statistical Analysis
System (Version 6.12, SAS Institute, Cary NC); to account for learning
effects we included order of treatment as a within-subject effect. Analysis of log-transformed mean power
output and performance time yielded percentage changes. We also used repeated measures analyses of
covariance to quantify the effect of individual differences in acclimation
(change in heart rate, perceived exertion, and hematocrit) on performance. Mean ±
standard deviation are used throughout for descriptive statistics. Precision of estimates of outcome
statistics is shown as the 95% confidence interval (CI), which defines the
likely range of the true value in the population. We used a spreadsheet (Hopkins, 2002) to calculate probabilities of
practical significance for the change in performance following heat
acclimation. The calculation requires
a value for the smallest worthwhile change in performance, which is ~0.5 of
an elite athlete's variation in performance time from race to race (Hopkins et al., 1999). For elite cyclists competing in
time trials lasting ~1 h, this variation is 1.3% (CD Paton and WG Hopkins,
unpublished observations), so the smallest worthwhile performance change is
~0.6%. Relative
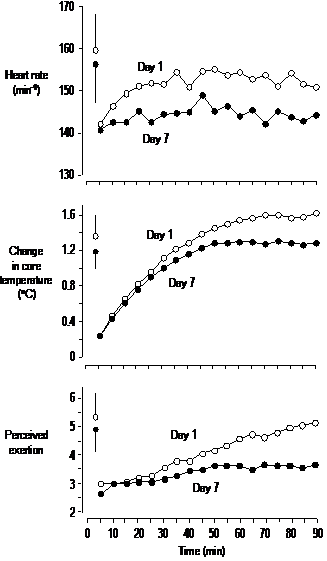
to training at normal temperature, training in the heat caused substantial
reductions in perceived exertion (1.5 units on the 10-point scale, CI = 0.3
to 2.8), core temperature (0.50 °C, CI = -0.16 to 1.17 °C), and heart rate (8.5 min-1, CI
= -2.5 to 19.5 min-1) during the standard exercise bout in the
heat relative to the same exercise in the thermoneutral condition (Figure
2). Training in the heat also caused a
slight decrease in sweat rate (77 ml·h-1, CI = -120 to 280 ml·h-1)
in the standard exercise bout, a modest decrease in hematocrit the day before
the performance test (-0.8%, CI = -2.0 to 0.4%), and a slight increase in
resting core temperature before the performance test (0.24 °C, CI = -0.09 to 0.57 °C). Change in
heart rate and change in perceived exertion were the most closely related of
the measures of acclimation (r = 0.93, CI = 0.70 to 0.99). Only three
subjects showed substantial changes in both these variables consistent with
acclimation (Figure 3). Change in core
temperature had only weak correlations with changes in heart rate (r = 0.15,
CI = -0.57 to 0.74), perceived exertion (r = -0.30, CI = -0.80 to 0.45), and
sweat rate (r = 0.33, CI = -0.43 to 0.82).
There was little relationship between change in sweat rate and changes
in resting core temperature, heart rate and perceived exertion (r = -0.36,
-0.15, -0.01). Change in hematocrit
had a strong correlation with change in sweat rate (r = -0.81, CI = -0.96 to
-0.32) but small-moderate correlations with change in perceived exertion,
heart rate, and core temperature (r = -0.14, -0.32, -0.53). Training
in the heat resulted in a slight enhancement of time-trial time from 54:36 ± 2:50 (min:s) before to 54:20 ± 3:30 after heat acclimation. After
adjustment for learning effects the enhancement was 0.4% (CI = -1.5 to
2.2%). The chances that the true
effect on performance is beneficial/trivial/harmful are 35/54/11%. Mean power
output in the time trial was 310 ± 40 W before heat acclimation and
315 ± 50 W after heat, an increase of
0.7% (CI = -4.0 to 5.3%). Training
in the heat had minimal effects on plasma-volume shift, heart rate, core
temperature, and sweat rate in the time trial (data not shown). There was also a minimal effect on arterial
oxygen saturation, but change in this variable correlated strongly with
change in hematocrit before the time trials
(r = 0.87, CI = 0.49 to 0.97) and change in sweat rate during the
acclimation rides (r = -0.81, CI = 0.32 to 0.96). Changes in other variables
measured during the time trial had generally trivial correlations with
measures of acclimation (data not shown).
A
substantial learning effect between the first and second time trials was
evident in performance time (1.7%, CI = -3.5 to 0.2%) and mean power output
(4.3%, CI = -0.3 to 8.9%). The
residual coefficient of variation in performance time was 1.6%, after learning
and treatment effects were accounted for in the statistical model. Individual Differences in Acclimation and Performance Changes in heart rate, perceived exertion, and hematocrit were
included in separate analyses of covariance to determine the extent to which
individual differences in acclimation predicted individual differences in
performance. For every 10 min-1
drop in heart rate after training in the heat, mean power declined by
1.3% (CI = -3.0% to 5.9%). Similarly,
for every unit decrease in perceived exertion, mean power also declined by
1.1% (CI = -2.7 to 5.0%). Figure 3
shows graphically how acclimation of heart rate and perceived exertion were
associated with these trends towards impairment of performance. In contrast, for every percent decrease in
hematocrit mean power improved by 1.0% (CI = -2.5% to 4.8%). DISCUSSION Changes
in heart rate, sweat rate, core temperature, perceived exertion, and plasma
volume have been used extensively to monitor heat acclimation in previous
studies (Armstrong and Maresh, 1991). In the present study there were substantial
changes in perceived exertion but only modest changes in heart rate, core
temperature, and sweat rate as a result of training in the heat. The 0.8% reduction in hematocrit with heat
acclimation is consistent with a modest
expansion of plasma volume. One
explanation for the apparently modest degree of heat acclimation could be
that most of our measures of heat acclimation were unreliable or otherwise
invalid. This explanation is not
consistent with the high correlation between some of these measures. The highest correlation, between changes in
heart rate and perceived exertion during the standard exercise bout,
indicates that both measures were probably tracking individual differences in
the extent of heat acclimation.
Similarly, the strong negative correlation between change in sweat
rate during the standard exercise bout and change in hematocrit is the
expected finding if there were individual differences in heat
acclimation. Curiously, these two
pairs of measures did not correlate well between each other, suggesting that
there may have been two largely independent adaptation processes in our
subjects. It is also possible that
changes in hematocrit were due to factors other than heat acclimation, such
as effects of diet or environment on hydration, which would also affect sweat
rate (Montain et al., 1995).
The one measure that probably was unreliable in our hands was change
in core temperature, which did not correlate well with any other
measure. The problem appears to have
been movement of the rectal thermister while the athletes were cycling. If we
make the reasonable assumption that changes in heart rate and perceived
exertion were valid and reliable measures of heat adaptation in our study, we
need to find explanations for the fact that the adaptation was only
modest. In most heat-acclimation
studies the subjects have been soldiers, mineworkers, or moderately trained
student volunteers (Maughan, 1997).
In contrast, our subjects were highly trained athletes. We therefore suspect that, for some of our
subjects, adaptations induced by hard training prior to the study resulted in
little or no thermoregulatory adaptation to the training performed in the
heat. The lack of adaptation might
have been due to high core temperature experienced during hard training (Davies and Thompson, 1986) or a circulatory adaptation to
hard training, such as expanded blood volume (Convertino, 1991). An important consequence for future studies
of heat adaptation is that researchers should use higher temperatures or more
intense exercise regimes when the subjects are highly trained athletes. The
observed effect of heat acclimation on performance time at normal temperature
was a little less than the smallest worthwhile improvement in performance, so
it is almost equally possible that the true effect is trivial or beneficial.
Although the true effect is unlikely to be harmful, further research with a
large sample or a more reliable measure of performance is needed for more
definitive conclusions. The
individual differences in heat acclimation had small inconsistent effects on
mean power in the time trial: there were tendencies for small negative
effects on performance for acclimation of heart rate and perceived exertion,
and a tendency towards a small positive effect for acclimation of hematocrit. Again, there was considerable uncertainty
in the true values of these effects, but the lack of consistency argues for
little overall true effect of acclimation on performance. A greater degree of acclimation than we
achieved might reveal a more marked effect on performance. A stronger and
more consistent relationship between performance and objective measures of
acclimation would also increase confidence that any enhancement of
performance is not a placebo effect, which can be ~1% for the performance test
with cyclists similar to those in the present study (Clark et al., 2000).
We would expect to see enhancement of performance with more
acclimation, but it is possible that the extra heat stress will induce some
kind of neural fatigue that counteracts beneficial physiological adaptations. In
principle, the observed small effect of acclimation on performance could be a
consequence of low validity or reliability of the measure of
performance. We are reasonably
confident about the validity, inasmuch as cyclists used their own bikes on
the Kingcycle ergometer, rode at a steady pace for most of the time trial,
and reached maximal or near maximal effort at the finish. The reliability of the measure (1.6% for
performance time) is somewhat larger than the 1.1% found by Palmer et al. (Palmer et al., 1996) in a reliability study of the
40-km time trial on the Kingcycle.
This difference is probably due to the learning effect between the
first and second time trial in our study.
The learning effect was large, probably because subjects were able to
see elapsed time and distance during the time trials and tried to improve
their performance in the second time trial.
Learning effects can reduce the reliability of performance tests,
because any individual differences in the learning effect add to
within-subject variability (Hopkins, 2000). In future studies of endurance
performance we intend to use a constant-load test to exhaustion, which
appears to be substantially more reliable than any other tests (Hopkins et al., 2001).
We will also blind the subjects to their times to exhaustion, to
reduce learning effects. In
conclusion, well-trained cyclists show only small adaptations to a moderate
heat-acclimation program. These
adaptations probably have little impact on performance at normal
environmental temperatures, but more research is needed to clarify this
issue. The
authors thank Diana Wilson and Frances Van Eerten for their valuable
technical assistance. Download these references as an Endnote file. Armstrong CG, Kenney WL (1993).
Effects of age and acclimation on responses to passive heat exposure. Journal
of Applied Physiology 75, 2162-2167 Armstrong
LE, Maresh CM (1991). The induction and decay of heat acclimatisation in
trained athletes. Sports Medicine 12, 302-312 Buono MJ,
Heaney JH, Canine KM (1998). Acclimation to humid heat lowers resting core
temperature. American Journal of Physiology 274, R1295-R1299 Caird SJ,
McKenzie AD, Sleivert GG (1999). Biofeedback and relaxation techniques
improve running economy in sub-elite long distance runners. Medicine and
Science in Sports and Exercise 31, 717-722 Clark VR,
Hopkins WG, Hawley JA, Burke LM (2000). Placebo effect of carbohydrate
feedings during a 40-km cycling time trial. Medicine and Science in Sports
and Exercise 32, 1642-1647 Convertino
VA (1991). Blood volume: its adaptation to endurance training. Medicine and
Science in Sports and Exercise 23, 1338-1348 Davies
CTM, Thompson MW (1986). Physiological responses to prolonged exercise in
ultramarathon athletes. Journal of Applied Physiology 61, 611-617 Greenleaf
JE, Convertino VA, Mangseth GR (1979). Plasma volume during stress in
man: osmolality and red cell volume.
Journal of Applied Physiology 47, 1031-1038 Hopkins
WG (2000). Measures of reliability in sports medicine and science. Sports
Medicine 30, 1-15 Hopkins
WG (2002). Calculating likely (confidence) limits and likelihoods for true
values (Excel spreadsheet). In: A new view of statistics. sportsci.org: Internet
Society for Sport Science, sportsci.org/resource/stats/xcl.xls Hopkins
WG, Hawley JA, Burke LM (1999). Design and analysis of research on sport
performance enhancement. Medicine and Science in Sports and Exercise 31,
472-485 Hopkins
WG, Schabort EJ, Hawley AJ (2001). Reliability of power in physical
performance tests. Sports Medicine 31, 211-234 Kirby CR,
Convertino VA (1986). Plasma aldosterone and sweat sodium concentrations after exercise and
heat acclimation. Journal of Applied Physiology 1986, 967-970 Kirwan
JP, Costill DL, Kuipers H, Burrell MJ, Fink WJ, Kovaleski JE, Fielding RA
(1987). Substrate utilisation in leg muscle of men after heat acclimation.
Journal of Applied Physiology 63, 31-35 Maughan R
(1997). Heat acclimatisation and rehydration strategy. British Journal of
Sports Medicine 31, 77 Montain
SJ, Latzka WA, Sawka MN (1995). Control of thermoregulatory sweating is
altered by hydration level and exercise intensity. Journal of Applied
Physiology 79, 1434-1439 Nadel ER,
Pandolf KB, Roberts MF, Stolwijk JAJ (1974). Mechanisms of thermal
acclimation to exercise and heat. Journal of Applied Physiology 37, 515-520 Noble B,
Borg GAV, Jacobs I, Ceci R, Kaiser P (1983). A category-ratio perceived
exertion scale: relationship to blood
lactate and heart rate. Medicine and Science in Sports and Exercise 15,
523-528 Palmer
GS, Dennis SC, Noakes TD, Hawley JA (1996). Assessment of the reproducability
of performance testing on an air-braked cycle ergometer. International
Journal of Sports Medicine 17, 293-298 Sawka MN,
Toner MM, Francesconi RP, Pandolf KB (1983). Hypohydration and exercise: effects of heat acclimation, gender, and
environment. Journal of Applied Physiology 55, 1147-1153 Senay LC,
Mitchell D, Wyndham CH (1976). Acclimatization in a hot, humid environment:
body fluid adjustments. Journal of Applied Physiology 40, 786-796 Williams
CG, Wyndham CH, Morrison JF (1967). Rate of loss of acclimatization in summer
and winter. Journal of Applied Physiology 22, 21-26 Young AJ,
Sawka MN, Levine L, Cadarette BS, Pandolf KB (1985). Skeletal muscle
metabolism during exercise in influenced by heat acclimation. Journal of
Applied Physiology 59, 1929-1935 |
|||||||||||||||||||||||||||||||||||||||||||